CSF is derived from blood plasma and is largely similar to it, except that CSF is nearly protein-free compared with plasma and has some different electrolyte levels. Due to the way it is produced, CSF has a higher chloride level than plasma, and an equivalent sodium level.[2][5]

What does cerebrospinal fluid consist of?
The electrolytes found in cerebrospinal fluid are sodium (Na+), chloride (Cl-), glucose, potassium (K+), calcium (Ca++), magnesium (Mg+), and . These are needed for osmoregulation, transport, neurotransmitter function, and conduction within the brain and spine. There are also other compounds present like enzymes, neurotransmitters and .
Summary The cerebrospinal fluid (CSF) is contained in the brain ventricles and the cranial and spinal subarachnoid spaces. The mean CSF volume is 150 ml, with 25 ml in the ventricles and 125 ml in subarachnoid spaces. CSF is predominantly, but not exclusively, secreted by the choroid plexuses. Brain interstitial fluid, ependyma and capillaries may also play a poorly defined role in CSF secretion. CSF circulation from sites of secretion to sites of absorption largely depends on the arterial pulse wave. Additional factors such as respiratory waves, the subject’s posture, jugular venous pressure and physical effort also modulate CSF flow dynamics and pressure. Cranial and spinal arachnoid villi have been considered for a long time to be the predominant sites of CSF absorption into the venous outflow system. Experimental data suggest that cranial and spinal nerve sheaths, the cribriform plate and the adventitia of cerebral arteries constitute substantial pathways of CSF drainage into the lymphatic outflow system. CSF is renewed about four times every 24 hours. Reduction of the CSF turnover rate during ageing leads to accumulation of catabolites in the brain and CSF that are also observed in certain neurodegenerative diseases. The CSF space is a dynamic pressure system. CSF pressure determines intracranial pressure with physiological values ranging between 3 and 4 mmHg before the age of one year, and between 10 and 15 mmHg in adults. Apart from its function of hydromechanical protection of the central nervous system, CSF also plays a prominent role in brain development and regulation of brain interstitial fluid homeostasis, which influences neuronal functioning. © 2011 Elsevier Masson SAS. All rights reserved
For a long time, the essential function of cerebrospinal fluid (CSF) was considered to be that of a fluid envelope that protects the central nervous system. Recent data derived from molecular biology show that CSF plays an essential role in homeostasis of the interstitial fluid of the brain parenchyma and regulation of neuronal functioning. Disorders of CSF hydrodynamics and composition are responsible for the major alterations of cerebral physiology observed in hydrocephalus and dementia, reflecting the importance of exchanges between CSF and the neuronal environment. Comparative anatomy Comparative anatomy of the meninges helps to elucidate the functional anatomy and ontogenesis of the CSF system in man [1]. The appearance of cerebrospinal fluid inside the neuraxis precedes circulation of cerebrospinal fluid in subarachnoid spaces during phylogenesis [2] The single primitive meninx with a large venous sinus in the spinal perimeningeal tissue of Selachii suggests the presence of a CSF venous absorption system. Large Teleostei present a pial layer lined by reticular tissue prefiguring the arachnoid membrane, but with no real CSF spaces. CSF is therefore contained in ventricular cavities. A peripheral fibrous layer differentiates and the perimeningeal tissue develops into an adipose tissue, which prefigures spinal epidural fat. In amphibians, reptiles and birds, the meninges comprise a dura mater and a pia mater. The perimeningeal tissue is considerably reduced, persisting at the spinal level in the form of epidural fat. In mammals, the subarachnoid space is clearly distinct from the pia mater. Participation of the central nervous system venous drainage in CSF absorption is first observed in Amniotes and is enhanced in the course of phylogenesis. Intracranial venous sinuses derived from cerebral epidural veins, and subarachnoid spaces develop in parallel [2]. Spinal epidural veins regress with a smaller participation in CSF absorption. The development of cerebrospinal fluid spaces retraces the steps of phylogenesis Cerebral and spinal meninges are derived from different embryonic tissues The three meningeal layers differentiate at the third month of intrauterine life. The meninges play a role in ontogenesis of the underlying brain tissue by inducing proliferation and differentiation of neuroblasts and axonal growth [3]. Experimental destruction of fetal meninges over the cerebellum induces cerebellar hypoplasia, neuronal ectopia and the formation of glial tissue in subarachnoid spaces [4,5]. Certain multiple malformation syndromes, such as Dandy Walker syndrome, comprising hypoplasia of the vermis and abnormalities of the cerebellar parenchyma and CSF spaces, could be due to similar mechanisms. The formation of subarachnoid spaces is not exclusively due to cerebrospinal fluid pressure On closure of the rostral and caudal neuropores at the first month of intrauterine life, the choroid plexuses are not yet functional [6]. However, CSF pressure increases in the lumen of the neural tube and the volume of the cephalic extremity increases, suggesting secretion of CSF by structures other than the choroid plexuses. The subarachnoid spaces appear on the 32nd day at the ventral aspect of the rhombencephalon, then extend caudally and dorsally However, the fourth ventricle is not yet open and CSF circulation is only effective on the 41st day. Formation of the subarachnoid spaces is therefore not exclusively due to CSF pressure. Formation of the subarachnoid spaces remains poorly understood. Capillaries appear to play a decisive role in the secretion and absorption of CSF during embryogenesis. Arachnoid cysts, dilatations of subarachnoid spaces predominantly located around blood vessels, appear to correspond to CSF spaces partly communicating with adjacent circulating blood sinuses. The first choroid plexuses The first choroid plexuses appear on the 41st day in the 4th ventricle [8]. The epithelium of the choroid plexus, continuous with the ependyma, is derived from the neural tube, while the leptomeningeal axis is derived from the paraxial mesoderm. The time at which the choroid plexuses start to secrete CSF has not been clearly determined. Arachnoid villi develop from the wall of intracranial venous sinuses From the 26th week, cerebral veins dilate at their anastomosis in the superior sagittal sinus. Villi are formed at the 35th week: the arachnoid stroma lined by endothelium protrudes into the lumen of the superior sagittal sinus via a defect in the dura mater. Real arachnoid granulations appear at the 39th week [9] and continue to develop until the age of about 18 months [10,11]. Cranial arachnoid granulations are essentially situated in contact with the posterior half of the superior sagittal sinus and adjacent venous lacunae and more rarely in contact with the transverse, superior petrosal, cavernous and sphenoparietal sinuses. These granulations ensure the bulk of CSF absorption at the end of organogenesis. However, comparative anatomy suggests other sites of CSF absorption in the absence of arachnoid villi or granulations. Volumes The CSF volume, estimated to be about 150 ml in adults, is distributed between 125 ml in cranial and spinal subarachnoid spaces and 25 ml in the ventricles, but with marked interindividual variations. Abnormally narrow ventricles, described as ‘‘slit ventricles’’, are observed in complex disorders of CSF circulation associated with cerebral oedema in patients with a CSF shunt. Inversely, hydrocephalus corresponds to an increased intracranial fluid volume and can be difficult to distinguish from cerebral atrophy, in which passive expansion of CSF spaces compensates for the reduction of brain volume. The distribution of fluid overload depends on the site of obstruction. In obstructive hydrocephalus, the obstruction is situated in the ventricular system, while in communicating hydrocephalus, the ventricular system and subarachnoid spaces freely communicate. The mechanisms of ventricular dilatation remain hypothetical, but include hydrodynamic factors (secretion and absorption rates, fluid pressure and cerebral compliance), hormonal neuropeptides, Atrial Natriuretic Peptide (ANP), and prostaglandin F2 (PGF2) [12]. Cerebrospinal fluid secretion Cerebrospinal fluid secretion in adults CSF secretion in adults varies between 400 to 600 ml per day, depending on the subject and the method used to study CSF secretion. Sixty to seventy-five percent of CSF is produced by the choroid plexuses of the lateral ventricles and the tela choroidea of the third and fourth ventricles. The choroid plexuses consist of granular meningeal protrusions into the ventricular lumen, the epithelial surface of which is continuous with the ependyma. They comprise a tuft of fenestrated capillaries. Choroidal cells present microvilli at their apical pole and are interconnected by tight junctions with a variable distribution according to the site on the ventricular wall [13]. Choroidal secretion of cerebrospinal fluid comprises two steps The first step consists of passive filtration of plasma from choroidal capillaries to the choroidal interstitial compartment according to a pressure gradient. The second step consists of active transport from the interstitial compartment to the ventricular lumen across the choroidal epithelium, involving carbonic anhydrase and membrane ion carrier proteins. Cytoplasmic carbonic anhydrase catalyses the formation of H+ and HCO3 − ions from water and CO2. The carrier proteins of basolateral membranes of choroidal cells exchange H+ and HCO3 − ions for Na+ and Cl− ions. ATP-dependent ion pumps of the apical membrane expel Na+, Cl−, HCO3 − and K+ ions towards the ventricular lumen. Water transport, facilitated by aquaporins I of the apical membrane, follows the osmotic gradients generated by these pumps [14]. The NaK2Cl cotransporter of the apical membrane generates ion transport in both directions and participates in regulation of CSF secretion and composition. Choroid plexuses secrete growth factors that probably act on the subventricular zone, which could repair tissue changes related to hydrocephalus, for example. They secrete vitamins B1, B12, C, folate, 2-microglobulin, arginine vasopressin and NO. Twenty percent of the peptides of CSF are derived from the brain and their concentration decreases as CSF flows from the ventricles to the subarachnoid spaces [15]. Extrachoroidal secretion Extrachoroidal secretion is derived from extracellular fluid and cerebral capillaries across the blood-brain barrier. This pathway appears to play a minimal role under physiological conditions. CSF can also be derived from the ependymal epithelium, the target of regulations mediated by neuropeptides and growth factors, which can be altered by ependymal changes induced, in particular, by ventricular dilatation. The composition of cerebrospinal fluid is not simply a plasma ultrafiltrate Na, Cl and Mg concentrations are higher and K and Ca concentrations are lower than those of plasma. The CSF cell count usually does not exceed five cells per milliliter. Variations in the closely regulated composition of CSF can be used for diagnostic purposes. Studies have demonstrated the existence of chronobiological cycles of Na content with peaks Na concentrations at 8:00 a.m. and at 6:00 p.m., with no modification of K and osmolarity. A relationship between the Na concentration and migraine has been proposed, as these peaks appear to correspond to the timing of migraine attacks [16]. Cerebrospinal fluid secretion and composition are finely regulated An increase in intraventricular pressure decreases the pressure gradient across the blood-brain barrier and decreases plasma filtration, but the capacities of adaptation of CSF secretion to intraventricular pressure at the initiation phase of hydrocephalus are rapidly exceeded. The choroid plexuses receive cholinergic, adrenergic, serotoninergic and peptidergic autonomic innervation. The sympathetic nervous system reduces CSF secretion, while the cholinergic system increases CSF secretion. The autonomic nervous system could be responsible for circadian variations of CSF secretion. Enzymes and membrane transporters are the targets of humoral regulation. Acid-base disorders modify the activity of carbonic anhydrase, aquaporins and membrane carrier proteins such as the NaK2Cl cotransporter. Monoamines and neuropeptide factors have also been shown to play a role. Dopamine, serotonin, melatonin, Atrial Natriuretic Peptide (ANP) and Arginine Vasopressin (AVP) receptors are present on the surface of choroidal epithelium. ANP and AVP decrease CSF secretion [17], as ANP acts on aquaporin I. The variable expression of ANP and AVP receptors according to CSF dynamics appears to be involved in the pathophysiology of hydrocephalus and dementia of the Alzheimer type. Loop diuretics and carbonic anhydrase inhibitors, which act on enzymatic mechanisms to decrease CSF secretion and turnover, could alter the neuronal environment, predisposing to age-related neurodegenerative disorders in the elderly. Cerebrospinal fluid circulation CSF circulation is a dynamic phenomenon and regulation of CSF circulation is responsible for cerebral homeostasis. CSF circulates from the sites of secretion to the sites of absorption according to a unidirectional rostrocaudal flow in ventricular cavities and a multidirectional flow in subarachnoid spaces. CSF flow is pulsatile, corresponding to the systolic pulse wave in choroidal arteries. CSF produced by the choroid plexuses in the lateral ventricles travels through interventricular foramina to the third ventricle, and then the fourth ventricle via the cerebral aqueduct and finally to the subarachnoid spaces via the median aperture (foramen of Magendie) of the fourth ventricle. In the cranial subarachnoid space, CSF circulates rostrally to the villous sites of absorption or caudally to the spinal subarachnoid space. Experimental studies have demonstrated the existence of a communication between CSF spaces and the adventitia of cerebral arteries: red blood cells injected into CSF spaces in the cat pass through the adventitia of cerebral arteries and are then detected in cervical lymph nodes [18]. The CSF, partly absorbed by spinal arachnoid villi, circulates rostrally to the cranial subarachnoid space. CSF flow is generated by the systolic pulse wave and rapid respiratory waves. The subcommissural organ, a differentiation of the ependyma at the rostral extremity of the cerebral aqueduct, appears to play a role in cerebrospinal fluid circulation The subcommissural organ synthesizes SCO-spondin, which has a phylogenetically conserved amino acid sequence [19]. SCO-spondin aggregates to form Reissner fibres, which guide the CSF circulation through the cerebral aqueduct. Rats immunised against Reissner fibres develop hydrocephalus due to stenosis of the cerebral aqueduct [20]. The subcommissural organ disappears early during development in man. An intrauterine abnormality of the subcommissural organ could explain certain forms of congenital hydrocephalus [21]. Cerebrospinal fluid absorption Cerebrospinal fluid is essentially absorbed into the internal jugular system via cranial arachnoid granulations Arachnoid villi are finger-like endothelium-lined protrusions of the arachnoid outer layer through the dura mater in the lumen of venous sinuses [22] (Fig. 1). Obstruction to internal jugular venous drainage is a rare cause of hydrocephalus. The pressure gradient between subarachnoid spaces and the venous sinus necessary to ensure CSF drainage is between 3 and 5 mmHg [23]. The pressure in the superior sagittal sinus remains relatively constant when the CSF pressure is modified experimentally [24]. Spinal arachnoid villi in contact with the epidural venous plexus represent a pathway of CSF absorption especially during effort (Fig. 2). Several different morphological types of arachnoid villi are present in the meningeal sheath of spinal nerve roots: some villi partially cross and others completely cross the dural membranes with various surface areas of exchange according to the degree of plication of the arachnoid layer. In the Green Monkey, arachnoid villi reach the epidural space and penetrate into the wall of veins situated around the spinal ganglion in about 16% of spinal roots [25]. Villous absorption of CSF, either in the brain or in the spine, is a dynamic process which adapts the filtration rate to CSF pressure (Fig. 1). In man, arachnoid villi in lumbosacral nerve roots increase CSF absorption in the upright position in response to gravity, and the absorbed CSF then enters the lymphatic system The role of extra-arachnoid absorption pathways remains poorly elucidated CSF can also be absorbed by cranial and spinal nerve sheaths, the ependyma and extracellular fluid according to pressure gradients. Absorption towards the interstitial compartment occurs via Virchow-Robin perivascular spaces. Cerebrospinal fluid absorption surfaces have been identified on meningeal sheaths CSF absorption surfaces have been identified on meningeal sheaths, particularly the meningeal recesses of spinal and cranial nerve roots, especially the trigeminal nerve and cochlear nerve. The optic nerve, derived from the diencephalon, presents a long extracranial course in its meningeal sheath. CISS MRI sequences of the orbits for assessment of hydrocephalus and benign intracranial hypertension show fluid thickening of the optic nerve sheaths, visualised as a high-intensity ring around the nerve, suggesting participation in CSF absorption when the capacity of the usual circulation-absorption pathways has been exceeded. The cribriform plate of the ethmoid bone has been studied in particular detail Vital stains injected into CSF spaces are subsequently found in the nasal submucosa and cervical lymph nodes [27,28]. This absorption pathway participates in the elimination of proteins and red blood cells from the CSF in cats and rabbits [29,30] and could be involved in the immune defense mechanisms of the brain [31]. The lymphatic circulation would be a preferential pathway of absorption of cerebral interstitial fluid, successively involving perivascular spaces, the arachnoid sheath of olfactory nerve fibres through the cribriform plate, the nasal submucosa and cervical lymph nodes [32]. In sheep, occlusion of the cribriform plate of the ethmoid bone increases the intracranial pressure [33], and lymphatic absorption of CSF increases with increasing intracranial pressure. At normal intracranial pressure, 10% of cervical lymph is derived from CSF, but when intracranial pressure increases from 10 to 70 cm H2O, 80% of cervical lymph is derived from CSF and the cervical lymphatic Figure 3 Cerebrospinal fluid (CSF) ‘‘secretion-circulationabsorption’’ process. CSF is mainly secreted by the choroid plexus and, to a lesser extent, by the interstitial compartment. It circulates rostrocaudally inside the ventricles and drains into the cerebellomedullary cistern (cisterna magna) through the median aperture (foramen of Magendie) of the fourth ventricle. CSF circulates in cranial and spinal subarachnoid spaces. In the cranial subarachnoid space, CSF flows towards arachnoid villi in the wall of venous sinuses from which it is absorbed. Part of the CSF is absorbed by the olfactory mucosa and cranial nerve (optic, trigeminal, facial and vestibulocochlear nerves) sheaths and is drained by the lymphatic system. In the spinal subarachnoid space, the part of the CSF absorbed by the epidural venous plexus and spinal nerve sheaths enters the lymphatic system, while the remaining CSF circulates rostrally towards the cranial subarachnoid space. CSF communicates with interstitial fluid via Virchow-Robin perivascular spaces. flow rate is increased fourfold [34]. Ligation of cervical lymph vessels of the dog induces cerebral oedema [35]. The functional role of this lymphatic pathway in man remains unknown (Fig. 3). These sites of absorptions constitute accessory pathways when the capacities of cranial arachnoid villi are exceeded. They are especially active in neonates, as immature arachnoid villi only become fully functional after the age of 18 months, and in the elderly due to fibrous changes of arachnoid granulations. Recent anatomical studies have focused on the cochlear aqueduct The cochlear aqueduct, situated in the petrous part of the temporal bone, establishes a communication between the subarachnoid space of the posterior cranial fossa and the perilymphatic space of the cochlea. This communication, patent in 93% of cases [36], would explain the impact of intracranial pressure variations on cochlear function, such as tinnitus occurring at high altitude and after ventriculoperitoneal shunting. Raised intracranial pressure modifies the results of certain audiological tests [37]. Cerebrospinal fluid turnover rate CSF is renewed four to five times per 24 hours in young adults. Ageing is characterised by a relative increase of the CSF compartment with respect to the brain parenchyma due to cerebral atrophy and a reduction of CSF turnover to three times a day at the age of 77 years. Catabolites of neurotransmitters and beta-amyloid (A) accumulate in the interstitial compartment, Virchow-Robin perivascular spaces [38,39], choroidal epithelium and ependyma during ageing and also in patients with adult chronic hydrocephalus (ACH) and Alzheimer’s disease (AD). Forty percent of patients with ACH present histological lesions of AD [40]. Decreased CSF A levels have been reported after an internal CSF shunt [41]. Cerebrospinal fluid pressure Cerebrospinal fluid pressure, defined as the intracranial pressure in the prone position, is the result of a dynamic equilibrium between CSF secretion, absorption and resistance to flow. CSF pressure can be measured invasively by a pressure transducer placed in the brain parenchyma or connected to CSF spaces via an external lumbar drain or external ventricular drain. Non-invasive methods essentially consist of interpretation of vascular flow on Doppler ultrasound. A method currently under investigation records the electrophysiological activity of outer hair cells of the cochlea. CSF pressure transmitted via the cochlear aqueduct influences intralabyrinthine pressure and the electrophysiological activity of outer hair cells. Monitoring of outer hair cell activity can therefore be used to monitor intracranial pressure variations [42—44]. Physiological values of cerebrospinal fluid pressure Physiological values of CSF pressure vary according to individuals and study methods between 10 and 15 mmHg in adults and 3 and 4 mmHg in infants. Higher values correspond to intracranial hypertension. CSF pressure varies with the systolic pulse wave, respiratory cycle, abdominal pressure, jugular venous pressure, state of arousal, physical activity and posture. The mechanisms regulating cerebrospinal fluid pressure have not been fully elucidated Cerebrospinal fluid pressure is determined by parenchymal and venous pressures. The cranial content comprises three compartments: parenchymal, venous and CSF. Prior to fontanelle closure, i.e. ‘‘open fontanelles’’, the plasticity of the infant’s skull adapts to pressure increases by an increase in intracranial volume leading to macrocephaly. After closure of the fontanelles, the skull forms a rigid bone chamber. In the presence of an intracranial space-occupying lesion, compensatory reductions of blood volumes (especially venous) and CSF are more active than at the ‘‘open fontanelles’’ state. The capacity of the intracranial contents to adapt to volume changes can be assessed by measuring brain compliance, defined as the volume required to modify intracranial pressure. Brain compliance is measured during a perfusion test, which monitors the increase in intracranial pressure generated by perfusion of saline into the spinal subarachnoid space. Brain compliance is higher in women and varies with age. The volume required to induce a tenfold increase in intracranial pressure is 8 ml in neonates, 20 ml in 2-year-old children and 26 ml in adults. Calculation of brain compliance must take into account the brain volume, which is an average of 335 ml in neonates and 1,250 ml in young adults. CSF pressure is regulated at all levels of CSF hydrodynamics: secretion, circulation, absorption. Increased intraventricular pressure exerts negative feedback on choroidal secretion by decreasing the pressure gradient across the blood-CSF barrier and by decreasing cerebral perfusion pressure. Neuropeptides (ANP and AVP) also appear to be involved. The concentrations of these neuropeptides in CSF and expression of their receptors in the choroidal epithelium increase with increasing CSF pressure and in acute hydrocephalus [45,46]. ANP and AVP decrease choroidal secretion of CSF and induce dilatation of pial arteries, which tends to compensate for the reduction of cerebral perfusion pressure in acute hydrocephalus [47]. Cerebrospinal fluid homeostasis CSF exerts a well-known function: hydromechanical protection of the neuraxis. CSF plays an essential role in homeostasis of cerebral interstitial fluid and the neuronal environment by regulation of the electrolyte balance, circulation of active molecules, and elimination of catabolites. CSF transports the choroidal plexus secretion products to their sites of action. This mode of distribution by CSF circulation modulates the activity of certain regions of the brain by impregnation, while synaptic transmission produces more rapid changes of activities [48]. The wastes of brain metabolism, peroxidation products and glycosylated proteins, accumulate with age-related decreased CSF turnover. Disclosure of interest The authors declare that they have no conflicts of interest concerning this article.
Introduction
Cerebrospinal fluid (CSF) is an ultrafiltrate of plasma contained within the ventricles of the brain and the subarachnoid spaces of the cranium and spine.[1] It performs vital functions, including providing nourishment, waste removal, and protection to the brain.[2] Adult CSF volume is estimated to be 150 ml, with a distribution of 125 ml within the subarachnoid spaces and 25 ml within the ventricles. CSF is predominantly secreted by the choroid plexus with other sources playing a more poorly defined role. In the adult population, its secretion varies between individuals, usually ranging from 400 to 600 ml per day. The constant secretion of CSF contributes to complete CSF renewal four to five times per 24-hour period in the average young adult. The reduction of CSF turnover may contribute to the accumulation of metabolites seen in aging and neurodegenerative diseases. The composition of CSF is strictly regulated, and any variation can be useful for diagnostic purposes.[1]
Cellular Level
Seventy to eighty percent of CSF production is via a network of modified ependymal cells known as the choroid plexus (CP).[1] The CP is a highly specialized, simple, cuboidal epithelium continuous with ependymal cells lining the ventricles of the brain. This simple cuboidal epithelium surrounds clusters of fenestrated capillaries allowing for the filtration of plasma.[3] CP cells have dense microvilli present on their apical surface. They are interconnected via tight junctions, creating a blood-CSF barrier that helps control the composition of CSF.[1]
As there is no appreciable barrier between the CSF and the extracellular space of the brain (ECSB), the blood-CSF barrier also serves to regulate the environment of the brain.[2] Larger substances such as cells, protein, and glucose are not allowed passage, whereas ions and small molecules such as vitamins and nutrients can pass into the CSF relatively easily.[4] Water is allowed passage through the CP epithelium via epithelial AQP1 channels. Substances that may not pass through the blood-CSF barrier, but are needed by the brain can be actively synthesized by or actively transported through the CP epithelial cells into the CSF. A 5-mV lumen positive voltage potential is present across CP epithelial cell membranes. This electrical potential difference pulls sodium, chloride, and bicarbonate ions from the plasma into the CSF, creating an osmotic gradient which then drives the movement of water into the CSF.[3]
When compared to plasma, CSF has a higher concentration of sodium, chloride, and magnesium, but a lower concentration of potassium and calcium.[1] Unlike plasma, CSF has only trace amounts of cells, protein, and immunoglobulins.[2] No cells can pass through the blood-CSF barrier, although small numbers of white blood cells are usually introduced to the CSF indirectly. The normal cell count of CSF is generally lower than 5 cells/ml.[1] Despite changes in blood composition and flow, the composition of CSF is kept constant, which provides a stable intraventricular environment, critical for maintaining normal neuronal function.[3]
Function
CSF assists the brain by providing protection, nourishment, and waste removal. CSF provides hydromechanical protection of the neuroaxis through two mechanisms. First, CSF acts as a shock absorber, cushioning the brain against the skull. Second, CSF allows the brain and spinal cord to become buoyant, reducing the effective weight of the brain from its normal 1,500 grams to a much lesser 50 grams. The reduction in weight lessens the force applied to the brain parenchyma and cerebral vessels during mechanical injury. Another function of CSF is to maintain homeostasis of the interstitial fluid of the brain. A stable environment for brain parenchyma is imperative for maintaining normal neuronal function.
The major conduit of nutrient supply to the brain is the CP-CSF-ECSB nexus. Substrates needed by the brain are transported from the blood, through the CP, into the CSF, and then diffuse into the ECSB for transportation to their sites of action within the brain. CSF also assists in the removal of brain metabolism waste products, such as peroxidation products, glycosylated proteins, excess neurotransmitters, debris from the lining of the ventricles, bacteria, viruses, and otherwise unnecessary molecules. Accumulation of such unnecessary molecules, seen in aging and some neurodegenerative diseases, interferes with neuronal functioning of the brain. The disruption of cerebral physiology experienced with the disruption of the hydrodynamics or composition of CSF suggests the importance of CSF functioning.[1][2][3]
Mechanism
CSF is continuously secreted with an unchanging composition, functioning to maintain a stable environment within the brain.[3] CSF is propelled along the neuroaxis from the site of secretion to the site of absorption, mainly by the rhythmic systolic pulse wave within the choroidal arteries. Lesser determinants of CSF flow are frequency of respiration, posture, venous pressure of the jugular vein, the physical effort of the individual, and time of day.[2]
CSF is secreted by the CPs located within the ventricles of the brain, with the two lateral ventricles being the primary producers. CSF flows throughout the ventricular system unidirectionally in a rostral to caudal manner. CSF produced in the lateral ventricles travel through the interventricular foramina to the third ventricle, through the cerebral aqueduct to the fourth ventricle, and then through the median aperture (also known as the foramen of Magendie) into the subarachnoid space at the base of the brain. Once in the subarachnoid space, the CSF begins to have a gentle multidirectional flow that creates an equalization of composition throughout the CSF. The CSF flows over the surface of the brain and down the length of the spinal cord while in the subarachnoid space. It leaves the subarachnoid space through arachnoid villi found along the superior sagittal venous sinus, intracranial venous sinuses, and around the roots of spinal nerves.
Arachnoid villi are protrusions of arachnoid mater through the dura mater into the lumen of a venous sinus. A 3 to 5 mmHg pressure gradient between the subarachnoid space and venous sinus pulls CSF into the venous outflow system through the arachnoid villi that help in its absorption. CSF may also enter into the lymphatic system via the nasal cribriform plate or spinal nerve roots. The clearance of CSF is dependent upon the posture of the individual, pressure differentials, and pathophysiology.[1][2]
Related Testing
Lumbar puncture (LP), also known as a spinal tap, is a commonly performed invasive procedure in which CSF is removed from the subarachnoid space. LP is used in the measurement of intracranial pressure and the sampling of CSF. It is commonly indicated in the evaluation of acute headaches and infections of the central nervous system. During an LP, the patient is placed in the lateral recumbent position. A sterile spinal needle is then slowly inserted between vertebrae, usually at the level of L3/4 or L4/5, into the subarachnoid space. Needle insertion may be guided by fluoroscopy or ultrasound to improve success rates and reduce the incidence of trauma.
Once CSF begins to flow through the needle, it is collected serially into four sterile tubes. Once collected, CSF can be analyzed for abnormally present or elevated CSF components, aiding in diagnosis. For example, the presence of xanthochromia, a yellow-orange discoloration of CSF caused by red blood cell degeneration, indicates the possibility of a subarachnoid hemorrhage. Elevation in concentrations of immunoglobulins, termed oligoclonal bands, may indicate the presence of a systemic infection or an autoimmune disease.
Contraindications of LP include raised intracranial pressure, bleeding disorders, and local skin infection. The procedure is relatively safe with seldom serious complications. Complications of LP include infection, bleeding, radicular pain, or cerebral herniation. The most common complication is a post-LP headache with symptoms beginning within 24 hours of the procedure and often resolving by day 10.[5][6]
Clinical Significance
Hydrocephalus is a pathological condition in which CSF abnormally accumulates due to increased CSF production, blockage of flow, or decreased absorption. The ventricles distend to accommodate elevated CSF volumes, potentially causing damage to the brain by pressing its tissue against the boney skull. Hydrocephalus may be congenital or acquired. Blocked CSF flow throughout the ventricles is classified as non-communicating, or obstructive, hydrocephalus. The blockage is often a mass such as a tumor or an abscess located within a foramen. Because CSF secretion is constant, obstruction of flow will lead to CSF build up in front of the blockage. For example, stenosis of the cerebral aqueduct, one of the most common causes of obstructive hydrocephalus, leads to enlargement of both lateral ventricles as well as the third ventricle. If the flow of CSF becomes obstructed outside the ventricles, in either the subarachnoid space or site of absorption, it classifies as communicating, or non-obstructive, hydrocephalus.
Hydrocephalus can be caused by genetic defects, infections, bleeding in the brain, trauma, or CNS tumors. Symptoms include headaches, convulsions, nausea, vomiting, disturbances of vision, and mental deterioration. The diagnosis is determined through imaging techniques such as ultrasound, computed tomogram (CT), or magnetic resonance imaging (MRI) scans. The most common treatment is shunt insertion, which diverts CSF away from the ventricles to an area of the body where it can be absorbed into circulation. Endoscopic third ventriculostomy, a procedure in which a hole is created in the floor of the third ventricle allowing CSF to bypass an obstruction, and cauterization of CP sections which decreases CSF production are other treatment options. If left untreated, hydrocephalus yields a risk of cognitive disturbances, physical disturbances, and death.[7][8]
CSF Leak is a condition in which CSF is able to escape from the subarachnoid space through a hole in the surrounding dura. The volume of CSF lost in a leak varies, ranging from minute to very substantial amounts. If the loss of CSF is great enough, spontaneous intracranial hypotension (SIH) may occur. SIH most often presents with a positional headache caused by downward displacement of the brain due to loss of buoyancy previously provided by the CSF. Posterior neck stiffness, nausea, and vomiting are also common symptoms. The incidence of SIH is estimated to be 5/100,000 annually. Women are twice as likely to be affected and have a peak age at around 40 years.
Diagnosis is aided by typical MRI findings, such as an increase of intracranial venous volume, pituitary hyperemia, enhanced pachymeninges, and descent of the brain. Many cases of SIH resolve without treatment. Conservative approaches such as bed rest, hydration, and increased caffeine intake may also be effective; however, more drastic measures may be necessary. An epidural blood patch, where blood is injected into the spinal epidural space, may relieve CSF hypovolemic symptoms by replacing lost CSF volume with blood volume. Surgical repair of the CSF leak via a suture or metal aneurysm clip is relatively safe and effective in providing relief.[9]
Meningitis is a condition in which the coverings of the brain become inflamed. There are two classifications of meningitis: aseptic and bacterial. Aseptic meningitis can result from agents such as fungi, medications, and cancer metastasis, but viruses cause the majority of aseptic meningitis cases. Fever, nuchal rigidity, and photophobia are classically presenting symptoms. Diagnosis is via an analysis of CSF obtained through LP. Viral PCR analysis of CSF is helpful in diagnosing viral meningitis. Treatment is often supportive, controlling fever and pain. Bacterial meningitis has a much lower incidence than aseptic meningitis, but is more serious. However, the incidence of bacterial meningitis has substantially dropped due to routine vaccination.
Symptoms are similar to aseptic meningitis, but clinical presentation is more severe. Additional symptoms include altered mental status, seizures, and focal neurologic signs. Diagnosis is also possible via LP. CSF is usually cloudy in appearance with a low glucose level and potential positive gram stain and culture. Patients presumed to have bacterial meningitis should immediately receive broad-spectrum antibiotics to prevent clinical deterioration. After culture results return, the clinician can make adjustments to the antibiotics. Patients should additionally be admitted to the intensive care unit for close monitoring. Most patients with bacterial meningitis who receive appropriate treatment recover without complications.[10]
Subarachnoid Hemorrhage (SAH) is the leakage of blood into the subarachnoid space where it mixes with CSF. Trauma is the most common cause of SAH with 80% of nontraumatic SAHs resulting from aneurysm rupture. Other nontraumatic causes of SAH include arteriovenous malformations and vasculitis. Spontaneous SAH has a low incidence, with only 30,000 cases worldwide annually. Ninety-seven percent of patients with SAH present with a sudden onset headache, described as a thunderclap headache or the worst headache of the patient's life. Other symptoms include vomiting, seizures, loss of consciousness, and death. Non-contrast head CT is useful in diagnosis. CT has high sensitivity after hemorrhage, but sensitivity decreases as time passes. After a negative CT, an LP should follow to rule out SAH. An LP is positive when erythrocytes are present in tubes 1 and 4, or xanthochromia is visible. Management of SAH consists of reducing risks of re-bleeding and avoiding any secondary brain injuries.[11]
Pseudotumor Cerebri Syndrome (PTCS) is a rare medical condition in which intracranial pressure is raised without the occurrence of ventriculomegaly or intracranial masses. The pathogenesis is not well understood. The most widely accepted theory proposes decreased absorption of CSF at the arachnoid granulations or the olfactory lymphatics as the cause. This condition has an annual incidence rate of 0.9/100,000 in the general population. Before puberty, both females and males are equally affected, but after puberty, women are affected nine times more often.
PTCS most commonly affects obese women of childbearing age. Women between the ages of 20 to 44 years and who weigh 20% more than their ideal body weight have an incidence rate of 19.3/100,000. A spinal fluid examination is critical in the diagnosis of PTCS. CSF pressure is greater than 250 mm CSF in adults and 280 mm CSF in children. Adolescents are the accepted values for diagnosis of PTCS. Headache is the most common presenting symptom, though there are no specific distinguishing characteristics of a PTCS headache. Occasionally asymptomatic patients present with papilledema detected during routine eye exams. Pulsatile tinnitus, transient visual obscurations, visual field defects, and visual loss are other symptoms of PTCS.
Traditional therapy includes medications to decrease CSF secretion from the choroid plexus. Surgery is indicated for patients with worsening vision caused by papilledema. Surgical options include optic nerve sheath fenestration and CSF shunting. Most patients with PTCS have a good outcome, although a small percentage of patients continue to experience persistent headaches or blindness.[12]
Cerebrospinal fluid
| Cerebrospinal fluid | |
|---|---|
 The cerebrospinal fluid circulates in the subarachnoid space around the brain and spinal cord, and in the ventricles of the brain. | |
 Image showing the location of CSF highlighting the brain's ventricular system | |
| Details | |
| Identifiers | |
| Latin | liquor cerebrospinalis |
| Acronym(s) | CSF |
| MeSH | D002555 |
| TA98 | A14.1.01.203 |
| TA2 | 5388 |
| Anatomical terminology | |
Cerebrospinal fluid (CSF) is a clear, colorless body fluid found within the tissue that surrounds the brain and spinal cord of all vertebrates.
CSF is produced by specialised ependymal cells in the choroid plexus of the ventricles of the brain, and absorbed in the arachnoid granulations. There is about 125 mL of CSF at any one time, and about 500 mL is generated every day. CSF acts as a shock absorber, cushion or buffer, providing basic mechanical and immunological protection to the brain inside the skull. CSF also serves a vital function in the cerebral autoregulation of cerebral blood flow.
CSF occupies the subarachnoid space (between the arachnoid mater and the pia mater) and the ventricular system around and inside the brain and spinal cord. It fills the ventricles of the brain, cisterns, and sulci, as well as the central canal of the spinal cord. There is also a connection from the subarachnoid space to the bony labyrinth of the inner ear via the perilymphatic duct where the perilymph is continuous with the cerebrospinal fluid. The ependymal cells of the choroid plexus have multiple motile cilia on their apical surfaces that beat to move the CSF through the ventricles.
A sample of CSF can be taken from around the spinal cord via lumbar puncture. This can be used to test the intracranial pressure, as well as indicate diseases including infections of the brain or the surrounding meninges.
Although noted by Hippocrates, it was forgotten for centuries, though later was described in the 18th century by Emanuel Swedenborg. In 1914, Harvey Cushing demonstrated that CSF is secreted by the choroid plexus.
Structure[edit]
Circulation[edit]


There is about 125–150 mL of CSF at any one time.[1] This CSF circulates within the ventricular system of the brain. The ventricles are a series of cavities filled with CSF. The majority of CSF is produced from within the two lateral ventricles. From here, CSF passes through the interventricular foramina to the third ventricle, then the cerebral aqueduct to the fourth ventricle. From the fourth ventricle, the fluid passes into the subarachnoid space through four openings – the central canal of the spinal cord, the median aperture, and the two lateral apertures.[1] CSF is present within the subarachnoid space, which covers the brain and spinal cord, and stretches below the end of the spinal cord to the sacrum.[1][2] There is a connection from the subarachnoid space to the bony labyrinth of the inner ear making the cerebrospinal fluid continuous with the perilymph in 93% of people.[3]
CSF moves in a single outward direction from the ventricles, but multidirectionally in the subarachnoid space.[3] Fluid movement is pulsatile, matching the pressure waves generated in blood vessels by the beating of the heart.[3] Some authors dispute this, posing that there is no unidirectional CSF circulation, but cardiac cycle-dependent bi-directional systolic-diastolic to-and-from cranio-spinal CSF movements.[4]
Contents[edit]
CSF is derived from blood plasma and is largely similar to it, except that CSF is nearly protein-free compared with plasma and has some different electrolyte levels. Due to the way it is produced, CSF has a higher chloride level than plasma, and an equivalent sodium level.[2][5]
CSF contains approximately 0.3% plasma proteins, or approximately 15 to 40 mg/dL, depending on sampling site.[6] In general, globular proteins and albumin are in lower concentration in ventricular CSF compared to lumbar or cisternal fluid.[7] This continuous flow into the venous system dilutes the concentration of larger, lipid-insoluble molecules penetrating the brain and CSF.[8] CSF is normally free of red blood cells and at most contains fewer than 5 white blood cells per mm3 (if the white cell count is higher than this it constitutes pleocytosis and can indicate inflammation or infection).[9]
Development[edit]
At around the third week of development, the embryo is a three-layered disc, covered with ectoderm, mesoderm and endoderm. A tube-like formation develops in the midline, called the notochord. The notochord releases extracellular molecules that affect the transformation of the overlying ectoderm into nervous tissue.[10] The neural tube, forming from the ectoderm, contains CSF prior to the development of the choroid plexuses.[3] The open neuropores of the neural tube close after the first month of development, and CSF pressure gradually increases.[3]
As the brain develops, by the fourth week of embryological development three swellings have formed within the embryo around the canal, near to where the head will develop. These swellings represent different components of the central nervous system: the prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain).[10] Subarachnoid spaces are first evident around the 32nd day of development near the rhombencephalon; circulation is visible from the 41st day.[3] At this time, the first choroid plexus can be seen, found in the fourth ventricle, although the time at which they first secrete CSF is not yet known.[3]
The developing forebrain surrounds the neural cord. As the forebrain develops, the neural cord within it becomes a ventricle, ultimately forming the lateral ventricles. Along the inner surface of both ventricles, the ventricular wall remains thin, and a choroid plexus develops, producing and releasing CSF.[10] CSF quickly fills the neural canal.[10] Arachnoid villi are formed around the 35th week of development, with arachnoid granulations noted around the 39th, and continuing developing until 18 months of age.[3]
The subcommissural organ secretes SCO-spondin, which forms Reissner's fiber within CSF assisting movement through the cerebral aqueduct. It is present in early intrauterine life but disappears during early development.[3]
Physiology[edit]
Function[edit]
CSF serves several purposes:
- Buoyancy: The actual mass of the human brain is about 1400–1500 grams; however, the net weight of the brain suspended in CSF is equivalent to a mass of 25-50 grams.[11][1] The brain therefore exists in neutral buoyancy, which allows the brain to maintain its density without being impaired by its own weight, which would cut off blood supply and kill neurons in the lower sections without CSF.[5]
- Protection: CSF protects the brain tissue from injury when jolted or hit, by providing a fluid buffer that acts as a shock absorber from some forms of mechanical injury.[1][5]
- Prevention of brain ischemia: The prevention of brain ischemia is aided by decreasing the amount of CSF in the limited space inside the skull. This decreases total intracranial pressure and facilitates blood perfusion.[1]
- Regulation: CSF allows for the homeostatic regulation of the distribution of substances between cells of the brain,[3] and neuroendocrine factors, to which slight changes can cause problems or damage to the nervous system. For example, high glycine concentration disrupts temperature and blood pressure control, and high CSF pH causes dizziness and fainting.[5]
- Clearing waste: CSF allows for the removal of waste products from the brain,[1] and is critical in the brain's lymphatic system, called the glymphatic system.[12] Metabolic waste products diffuse rapidly into CSF and are removed into the bloodstream as CSF is absorbed.[13] When this goes awry, CSF can become toxic, such as in amyotrophic lateral sclerosis, the commonest form of motor neuron disease.[14][15]
Production[edit]
| Substance | CSF | Serum |
|---|---|---|
| Water content (% wt) | 99 | 93 |
| Protein (mg/dL) | 35 | 7000 |
| Glucose (mg/dL) | 60 | 90 |
| Osmolarity (mOsm/L) | 295 | 295 |
| Sodium (mEq/L) | 138 | 138 |
| Potassium (mEq/L) | 2.8 | 4.5 |
| Calcium (mEq/L) | 2.1 | 4.8 |
| Magnesium (mEq/L) | 2.0–2.5[16] | 1.7 |
| Chloride (mEq/L) | 119 | 102 |
| pH | 7.33 | 7.41 |
The brain produces roughly 500 mL of cerebrospinal fluid per day at a rate of about 20 mL an hour.[17] This transcellular fluid is constantly reabsorbed, so that only 125–150 mL is present at any one time.[1]
CSF volume is higher on a mL per kg body weight basis in children compared to adults. Infants have a CSF volume of 4 mL/kg, children have a CSF volume of 3 mL/kg, and adults have a CSF volume of 1.5–2 mL/kg. A high CSF volume is why a larger dose of local anesthetic, on a mL/kg basis, is needed in infants.[citation needed] Additionally, the larger CSF volume may be one reason as to why children have lower rates of postdural puncture headache.[18]
Most (about two-thirds to 80%) of CSF is produced by the choroid plexus.[1][2] The choroid plexus is a network of blood vessels present within sections of the four ventricles of the brain. It is present throughout the ventricular system except for the cerebral aqueduct, and the frontal and occipital horns of the lateral ventricles.[19] CSF is mostly produced by the lateral ventricles.[17] CSF is also produced by the single layer of column-shaped ependymal cells which line the ventricles; by the lining surrounding the subarachnoid space; and a small amount directly from the tiny spaces surrounding blood vessels around the brain.[2]
CSF is produced by the choroid plexus in two steps. Firstly, a filtered form of plasma moves from fenestrated capillaries in the choroid plexus into an interstitial space,[1] with movement guided by a difference in pressure between the blood in the capillaries and the interstitial fluid.[3] This fluid then needs to pass through the epithelium cells lining the choroid plexus into the ventricles, an active process requiring the transport of sodium, potassium and chloride that draws water into CSF by creating osmotic pressure.[3] Unlike blood passing from the capillaries into the choroid plexus, the epithelial cells lining the choroid plexus contain tight junctions between cells, which act to prevent most substances flowing freely into CSF.[20] Cilia on the apical surfaces of the ependymal cells beat to help transport the CSF.[21]
Water and carbon dioxide from the interstitial fluid diffuse into the epithelial cells. Within these cells, carbonic anhydrase converts the substances into bicarbonate and hydrogen ions. These are exchanged for sodium and chloride on the cell surface facing the interstitium.[3] Sodium, chloride, bicarbonate and potassium are then actively secreted into the ventricular lumen.[2][3] This creates osmotic pressure and draws water into CSF,[2] facilitated by aquaporins.[3] CSF contains many fewer protein anions than blood plasma. Protein in the blood is primarily composed of anions where each anion has many negative charges on it.[22]
As a result, to maintain electroneutrality blood plasma has a much lower concentration of chloride anions than sodium cations. CSF contains a similar concentration of sodium ions to blood plasma but fewer protein cations and therefore a smaller imbalance between sodium and chloride resulting in a higher concentration of chloride ions than plasma. This creates an osmotic pressure difference with the plasma. CSF has less potassium, calcium, glucose and protein.[5] Choroid plexuses also secrete growth factors, iodine,[23] vitamins B1, B12, C, folate, beta-2 microglobulin, arginine vasopressin and nitric oxide into CSF.[3] A Na-K-Cl cotransporter and Na/K ATPase found on the surface of the choroid endothelium, appears to play a role in regulating CSF secretion and composition.[3][1]
It has been hypothesised that CSF is not primarily produced by the choroid plexus, but is being permanently produced inside the entire CSF system, as a consequence of water filtration through the capillary walls into the interstitial fluid of the surrounding brain tissue, regulated by AQP-4.[4]
There are circadian variations in CSF secretion, with the mechanisms not fully understood, but potentially relating to differences in the activation of the autonomic nervous system over the course of the day.[3]
Choroid plexus of the lateral ventricle produces CSF from the arterial blood provided by the anterior choroidal artery.[24] In the fourth ventricle, CSF is produced from the arterial blood from the anterior inferior cerebellar artery (cerebellopontine angle and the adjacent part of the lateral recess), the posterior inferior cerebellar artery (roof and median opening), and the superior cerebellar artery.[25]
Reabsorption[edit]
CSF returns to the vascular system by entering the dural venous sinuses via arachnoid granulations.[2] These are outpouchings of the arachnoid mater into the venous sinuses around the brain, with valves to ensure one-way drainage.[2] This occurs because of a pressure difference between the arachnoid mater and venous sinuses.[3] CSF has also been seen to drain into lymphatic vessels,[26] particularly those surrounding the nose via drainage along the olfactory nerve through the cribriform plate. The pathway and extent are currently not known,[1] but may involve CSF flow along some cranial nerves and be more prominent in the neonate.[3] CSF turns over at a rate of three to four times a day.[2] CSF has also been seen to be reabsorbed through the sheathes of cranial and spinal nerve sheathes, and through the ependyma.[3]
Regulation[edit]
The composition and rate of CSF generation are influenced by hormones and the content and pressure of blood and CSF.[3] For example, when CSF pressure is higher, there is less of a pressure difference between the capillary blood in choroid plexuses and CSF, decreasing the rate at which fluids move into the choroid plexus and CSF generation.[3] The autonomic nervous system influences choroid plexus CSF secretion, with activation of the sympathetic nervous system decreasing secretion and the parasympathetic nervous system increasing it.[3] Changes in the pH of the blood can affect the activity of carbonic anhydrase, and some drugs (such as furosemide, acting on the Na-Cl cotransporter) have the potential to impact membrane channels.[3]
Clinical significance[edit]
Pressure[edit]
CSF pressure, as measured by lumbar puncture, is 10–18 cmH2O (8–15 mmHg or 1.1–2 kPa) with the patient lying on the side and 20–30 cmH2O (16–24 mmHg or 2.1–3.2 kPa) with the patient sitting up.[27] In newborns, CSF pressure ranges from 8 to 10 cmH2O (4.4–7.3 mmHg or 0.78–0.98 kPa). Most variations are due to coughing or internal compression of jugular veins in the neck. When lying down, the CSF pressure as estimated by lumbar puncture is similar to the intracranial pressure.
Hydrocephalus is an abnormal accumulation of CSF in the ventricles of the brain.[28] Hydrocephalus can occur because of obstruction of the passage of CSF, such as from an infection, injury, mass, or congenital abnormality.[28][29] Hydrocephalus without obstruction associated with normal CSF pressure may also occur.[28] Symptoms can include problems with gait and coordination, urinary incontinence, nausea and vomiting, and progressively impaired cognition.[29] In infants, hydrocephalus can cause an enlarged head, as the bones of the skull have not yet fused, seizures, irritability and drowsiness.[29] A CT scan or MRI scan may reveal enlargement of one or both lateral ventricles, or causative masses or lesions,[28][29] and lumbar puncture may be used to demonstrate and in some circumstances relieve high intracranial pressure.[30] Hydrocephalus is usually treated through the insertion of a shunt, such as a ventriculo-peritoneal shunt, which diverts fluid to another part of the body.[28][29]
Idiopathic intracranial hypertension is a condition of unknown cause characterized by a rise in CSF pressure. It is associated with headaches, double vision, difficulties seeing, and a swollen optic disc.[28] It can occur in association with the use of vitamin A and tetracycline antibiotics, or without any identifiable cause at all, particularly in younger obese women.[28] Management may include ceasing any known causes, a carbonic anhydrase inhibitor such as acetazolamide, repeated drainage via lumbar puncture, or the insertion of a shunt such as a ventriculoperitoneal shunt.[28]
CSF leak[edit]
CSF can leak from the dura as a result of different causes such as physical trauma or a lumbar puncture, or from no known cause when it is termed a spontaneous cerebrospinal fluid leak.[31] It is usually associated with intracranial hypotension: low CSF pressure.[30] It can cause headaches, made worse by standing, moving and coughing,[30] as the low CSF pressure causes the brain to "sag" downwards and put pressure on its lower structures.[30] If a leak is identified, a beta-2 transferrin test of the leaking fluid, when positive, is highly specific and sensitive for the detection for CSF leakage.[31] Medical imaging such as CT scans and MRI scans can be used to investigate for a presumed CSF leak when no obvious leak is found but low CSF pressure is identified.[32] Caffeine, given either orally or intravenously, often offers symptomatic relief.[32] Treatment of an identified leak may include injection of a person's blood into the epidural space (an epidural blood patch), spinal surgery, or fibrin glue.[32]
Lumbar puncture[edit]

CSF can be tested for the diagnosis of a variety of neurological diseases, usually obtained by a procedure called lumbar puncture.[33] Lumbar puncture is carried out under sterile conditions by inserting a needle into the subarachnoid space, usually between the third and fourth lumbar vertebrae. CSF is extracted through the needle, and tested.[31] About one third of people experience a headache after lumbar puncture,[31] and pain or discomfort at the needle entry site is common. Rarer complications may include bruising, meningitis or ongoing post lumbar-puncture leakage of CSF.[1]
Testing often includes observing the colour of the fluid, measuring CSF pressure, and counting and identifying white and red blood cells within the fluid; measuring protein and glucose levels; and culturing the fluid.[31][33] The presence of red blood cells and xanthochromia may indicate subarachnoid hemorrhage; whereas central nervous system infections such as meningitis, may be indicated by elevated white blood cell levels.[33] A CSF culture may yield the microorganism that has caused the infection,[31] or PCR may be used to identify a viral cause.[33] Investigations to the total type and nature of proteins reveal point to specific diseases, including multiple sclerosis, paraneoplastic syndromes, systemic lupus erythematosus, neurosarcoidosis, cerebral angiitis;[1] and specific antibodies such as aquaporin-4 may be tested for to assist in the diagnosis of autoimmune conditions.[1] A lumbar puncture that drains CSF may also be used as part of treatment for some conditions, including idiopathic intracranial hypertension and normal pressure hydrocephalus.[1]
Lumbar puncture can also be performed to measure the intracranial pressure, which might be increased in certain types of hydrocephalus. However, a lumbar puncture should never be performed if increased intracranial pressure is suspected due to certain situations such as a tumour, because it can lead to fatal brain herniation.[31]
Anaesthesia and chemotherapy[edit]
Some anaesthetics and chemotherapy are injected intrathecally into the subarachnoid space, where they spread around CSF, meaning substances that cannot cross the blood–brain barrier can still be active throughout the central nervous system.[34][35] Baricity refers to the density of a substance compared to the density of human cerebrospinal fluid and is used in regional anesthesia to determine the manner in which a particular drug will spread in the intrathecal space.[34]
History[edit]
Various comments by ancient physicians have been read as referring to CSF. Hippocrates discussed "water" surrounding the brain when describing congenital hydrocephalus, and Galen referred to "excremental liquid" in the ventricles of the brain, which he believed was purged into the nose. But for some 16 intervening centuries of ongoing anatomical study, CSF remained unmentioned in the literature. This is perhaps because of the prevailing autopsy technique, which involved cutting off the head, thereby removing evidence of CSF before the brain was examined.[36]
The modern rediscovery of CSF is credited to Emanuel Swedenborg. In a manuscript written between 1741 and 1744, unpublished in his lifetime, Swedenborg referred to CSF as "spirituous lymph" secreted from the roof of the fourth ventricle down to the medulla oblongata and spinal cord. This manuscript was eventually published in translation in 1887.[36]
Albrecht von Haller, a Swiss physician and physiologist, made note in his 1747 book on physiology that the "water" in the brain was secreted into the ventricles and absorbed in the veins, and when secreted in excess, could lead to hydrocephalus.[36] François Magendie studied the properties of CSF by vivisection. He discovered the foramen Magendie, the opening in the roof of the fourth ventricle, but mistakenly believed that CSF was secreted by the pia mater.[36]
Thomas Willis (noted as the discoverer of the circle of Willis) made note of the fact that the consistency of CSF is altered in meningitis.[36] In 1869 Gustav Schwalbe proposed that CSF drainage could occur via lymphatic vessels.[1]
In 1891, W. Essex Wynter began treating tubercular meningitis by removing CSF from the subarachnoid space, and Heinrich Quincke began to popularize lumbar puncture, which he advocated for both diagnostic and therapeutic purposes.[36] In 1912, a neurologist William Mestrezat gave the first accurate description of the chemical composition of CSF.[36] In 1914, Harvey W. Cushing published conclusive evidence that CSF is secreted by the choroid plexus.[36]
Other animals[edit]
During phylogenesis, CSF is present within the neuraxis before it circulates.[3] The CSF of Teleostei fish, which do not have a subarachnoid space, is contained within the ventricles of their brains.[3] In mammals, where a subarachnoid space is present, CSF is present in it.[3] Absorption of CSF is seen in amniotes and more complex species, and as species become progressively more complex, the system of absorption becomes progressively more enhanced, and the role of spinal epidural veins in absorption plays a progressively smaller and smaller role.[3]
The amount of cerebrospinal fluid varies by size and species.[37] In humans and other mammals, cerebrospinal fluid turns over at a rate of 3–5 times a day.[37] Problems with CSF circulation, leading to hydrocephalus, can occur in other animals as well as humans.[37]
Cerebrospinal Fluid (CSF) Analysis





What is a Cerebrospinal Fluid (CSF) Analysis?
Cerebrospinal fluid (CSF) is a clear, colorless, watery fluid that flows in and around your brain and spinal cord. Your brain and spinal cord make up your central nervous system. It controls and coordinates everything you do, including your ability to move, breathe, see think, and more.
Cerebrospinal fluid acts like a cushion that helps protect your brain and spinal cord from sudden impact or injury. The fluid also removes waste products from the brain and helps your central nervous system work properly.
A CSF analysis is a group of tests that use a sample of your cerebrospinal fluid to help diagnose diseases of the brain and spinal cord and other conditions that affect the central nervous system.
Other names: Spinal Fluid Analysis, CSF Analysis
What is it used for?
A CSF analysis is used to measure different substances in your cerebrospinal fluid. It may include tests to diagnose:
- Infectious diseases of the brain and spinal cord, including meningitis and encephalitis. CSF tests for infections look at white blood cells, bacteria, and other substances in the cerebrospinal fluid
- Autoimmune disorders, such as Guillain-Barré Syndrome and multiple sclerosis (MS). CSF tests for these disorders look for high levels of certain proteins in the cerebrospinal fluid.
- Bleeding in the brain
- Brain tumors, including from cancers in other parts of the body that may spread to the central nervous system
- Alzheimer's disease, the most common form of dementia, which includes memory loss, confusion, and changes in behavior
Why do I need a CSF analysis?
Your health care provider may order a CSF analysis if you have:
- Symptoms of an infection or bleeding in the brain or spinal cord
- Symptoms of an autoimmune disorder, such as multiple sclerosis (MS)
- Had a brain injury or an injury to your spinal cord
- Have cancer that may have spread to your central nervous system
- Symptoms that may be from another central nervous system condition, such as headaches
Symptoms of a brain or spinal cord infection include:
- Fever
- Severe headache
- Seizures
- Stiff neck
- Nausea and vomiting
- Sensitivity to light
- Double vision
- Changes in behavior
- Confusion
Symptoms of MS often vary and come and go, or they may steadily get worse. They may include:
- Blurred or double vision
- Tingling, numbness, or pain in the arms, legs, body, or face
- Painful muscle spasms
- Weak muscles, often in the hands and legs
- Dizziness, balance problems, or clumsy movement when walking
- Bladder control problems
- Fatigue
- Problems thinking or learning new things
What happens during a CSF analysis?
To get a sample of cerebrospinal fluid, a provider will do a procedure called a spinal tap, also known as a lumbar puncture. A spinal tap is usually done in a hospital. During the procedure:
- You will lie on your side or sit on an exam table.
- A provider will clean your back and inject an anesthetic into your skin, so you won't feel pain during the procedure. Your provider may put a numbing cream on your back before this injection.
- When the area on your back is completely numb, your provider will insert a thin, hollow needle between two vertebrae in your lower spine. Vertebrae are the small backbones that make up your spine.
- Your provider will withdraw a small amount of cerebrospinal fluid for testing. This will take about five minutes.
- You'll need to stay very still while the fluid is being withdrawn.
- Your provider may ask you to lie on your back for an hour or two after the procedure. This may prevent you from getting a headache afterward.
Will I need to do anything to prepare for the test?
You don't need any special preparations for a CSF analysis, but you may be asked to empty your bladder (pee) and bowels (poop) before the test.
Are there any risks to the test?
There is very little risk to having a spinal tap. You may feel a little pinch or pressure when the needle is inserted. After the test, you may feel some pain or tenderness in your back at the site where the needle was inserted.
You may also have some bleeding at the site or get a headache. The headache may last for several hours or up to a week or more, but your provider may suggest treatment to help relieve the pain.
What do the results mean?
A CSF analysis may include a variety of different tests on your sample. So, the measurements on your test results will depend on which tests were done. Your provider can explain what your results mean.
In general, your CSF analysis results may show that you have an infection, an autoimmune disorder, such as multiple sclerosis (MS), or another disease or injury of the brain or spinal cord. Your provider will likely order more tests to confirm your diagnosis.
Learn more about laboratory tests, reference ranges, and understanding results.
Is there anything else I need to know about a CSF analysis?
Some infections, such as meningitis caused by bacteria, are life-threatening emergencies. If your provider suspects you have bacterial meningitis or another serious infection, you may need to start medicine before you have a final diagnosis.
Introduction
Cerebrospinal fluid (CSF) is a clear, plasma-like fluid (an ultrafiltrate of plasma) that bathes the central nervous system (CNS). It occupies the central spinal canal, the ventricular system, and the subarachnoid space.

Image 1 shows a 3D image of the CSF circulation as it would look inside the skull.
- CSF performs vital functions including: Support; Shock absorber; Homeostasis; Nutrition; Immune function.[1]
- Adult CSF volume is estimated to be 150 ml with a distribution of 125 ml within the subarachnoid spaces and 25 ml within the ventricles.
- Ependymal cells, which create cerebral spinal fluid (CSF), line the ventricles of the brain and central canal of the spinal cord
- CSF is predominantly secreted by the choroid plexus with other sources playing a more poorly defined role, an adult produces between 400 to 600 ml per day.
- The constant secretion of CSF contributes to complete CSF renewal four to five times per 24-hour period in the average young adult.
- Reduction of CSF turnover may contribute to the accumulation of metabolites seen in aging and neurodegenerative diseases. The composition of CSF is closely regulated, and any variation can be used for diagnostic purposes[2]
- Out-pouching of the arachnoid mater (arachnoid granulations) are responsible for the resorption of CSF into the dural venous sinuses.
- Disequilibrium in synthesis and resorption or obstruction of circulation results in CSF accumulation and raised intracranial pressure called hydrocephalus.
- CSF be examined clinically through a lumbar puncture. With a lumbar puncture, physicians can look for abnormalities in the CSF, which can be helpful when creating a differential diagnosis[3]
Structure and Function
- Support - The CSF supports the weight of the brain estimated at 1500 gm and suspends it in neutral buoyancy to a net weight of about 25 gm. Hence, the entire brain density is cushioned, protecting it from crushing into the bony cranium.
- Shock absorber - It protects the brain from damage during head trauma. Otherwise, even minor head bopping will result in severe brain injury.
- Homeostasis - The biochemical constituents and volume of the CSF play vital cerebral homeostatic roles:
- Maintains stable intrinsic CNS temperature
- Biochemical constituents and electrolytes maintain the osmotic pressure responsible for normal CSF pressure which is essential to maintaining normal cerebral perfusion
- Biochemical waste products diffuse into the CSF and are removed as CSF is resorbed through arachnoid granulations into venous circulation, a small percentage of CSF also drains into lymphatic circulation.
4. Nutrition - The CSF contains glucose, proteins, lipids, and electrolytes, providing essential CNS nutrition.
5. Immune function - The CSF contains immunoglobulins and mononuclear cells.[1]
The 5 minute video below is a great summary of the CSF
Ventricles and CSF
- CSF is produced mainly by a structure called the choroid plexus in the lateral, third and fourth ventricles.
- CSF flows from the lateral ventricle to the third ventricle through the interventricular foramen (also called the foramen of Monro).
- The third ventricle and fourth ventricle are connected to each other by the cerebral aqueduct (also called the Aqueduct of Sylvius).
- CSF then flows into the subarachnoid space through the foramina of Luschka (there are two of these) and the foramen of Magendie (only one of these).
- Absorption of the CSF into the blood stream takes place in the superior sagittal sinus through structures called arachnoid villi.
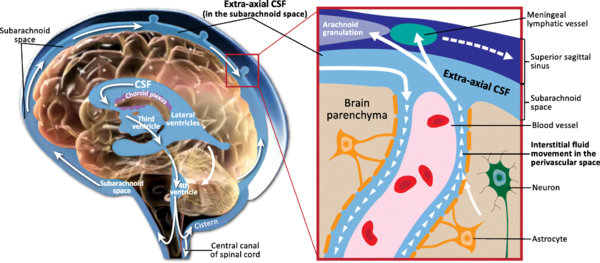

When the CSF pressure is greater than the venous pressure, CSF will flow into the blood stream. However, the arachnoid villi act as "one way valves"...if the CSF pressure is less than the venous pressure, the arachnoid villi will NOT let blood pass into the ventricular system[5]
Image 2 shows Schematic of CSF circulation, CSF outflow systems, and the anatomy of various CSF compartments.
Blood Supply and Lymphatics
The choroid plexus is a group of fenestrated blood capillaries located in the ventricular system. The choroid plexus mostly synthesizes CSF.
Arachnoid granulations are responsible for CSF resorption; they drain CSF into the dural venous sinuses.
CSF drains into the lymphatic circulation, via lymph ducts contiguous to the olfactory duct, as it passes through the cribriform plate.[1]
Clinical Significance
Hydrocephalus is a pathological condition of abnormal accumulation of CSF caused by increased CSF production, blockage of flow, or decreased absorption. The ventricles distend in order to accommodate elevated CSF volumes, potentially causing damage to the brain by pressing its tissue against the boney skull. Hydrocephalus may be congenital or acquired.[2]
CSF Leak is a condition in which CSF is able escape from the subarachnoid space through a hole in the surrounding dura. The volume of CSF lost in a leak is very variable, ranging from insignificant to very substantial amounts.
Syringomyelia due to blockage of CSF circulation.
Meningitis is a condition in which the coverings of the brain become inflamed. There are two classifications of meningitis: aseptic and bacterial. Aseptic meningitis can be caused by agents such as fungi, medications, and cancer metastasis, but the majority of aseptic meningitis cases are caused by viruses. Fever, nuchal rigidity, and photophobia are classic presenting symptoms. Diagnosis is made via an analysis of CSF obtained through LP.
Subarachnoid Hemorrhage (SAH) is the leakage of blood into the subarachnoid space where it mixes with the CSF. SAH is most commonly caused by trauma with 80% of nontraumatic SAHs being caused by aneurysm rupture. Other nontraumatic causes of SAH include arteriovenous malformations and vasculitis.[2]
Lumber Puncture and CSF Analysis - Lumbar puncture is a sterile procedure, done to obtain CSF samples for diagnostic purposes. It involves passing a needle into the subarachnoid space at the levels between L2 and L5 vertebrae. However, most commonly lumbar puncture is performed between L4 and L5. Biochemical, microbiologic and cytologic studies are then carried out on the sample.[1]
Conclusion
Cerebrospinal fluid (CSF) plays an essential role in maintaining the homeostasis of the central nervous system.
The functions of CSF include: (1) buoyancy of the brain, spinal cord, and nerves; (2) volume adjustment in the cranial cavity; (3) nutrient transport; (4) protein or peptide transport; (5) brain volume regulation through osmoregulation; (6) buffering effect against external forces; (7) signal transduction; (8) drug transport; (9) immune system control; (10) elimination of metabolites and unnecessary substances; and finally cooling of heat generated by neural activity.
For CSF to fulfil these functions, fluid-like movement in the ventricles and subarachnoid space is necessary.[6]
Abnormalities in the CSF flow or introduction of infection and or irritants can seriously affect an individuals functioning.
إرسال تعليق